Страница: 6/13
Смена растворов производилась протоком, скорость которого можно было варьировать от 10 до 20 мл/мин. Объем экспериментальной камеры составлял 0.5 мл. Все эксперименты были проведены при температуре (32 °С).
|
|

Аппликация АТФ в большинстве нейронов моторной коры (98 из 102) вызывает быстрое и кратковременное повышение концентрации цитозольного кальция [Ca2+]i . Концентрация [Ca2+]i достигает своего максимума на протяжении 6-8 секунд и затем восстанавливается до базального уровня при продолжающемся присутствии АТФ. Отмывание АТФ приводит к восстановлению [Ca2+]i до уровня покоя. Характерный вид АТФ - индуцированного кальциевого транзиента представлен на рисунке 5.
|
|
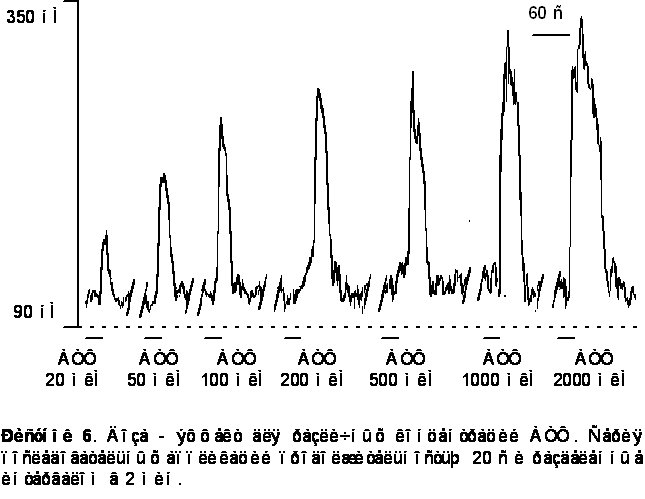
Амплитуда АТФ - индуцированного кальциевого транзиента варьировалась в пределах от 50 до 450 нМ. Среднее значение выброса [Ca2+]i вызванного приложением АТФ в концентрации 100 мкМ находилось в пределах 200-250 нМ.
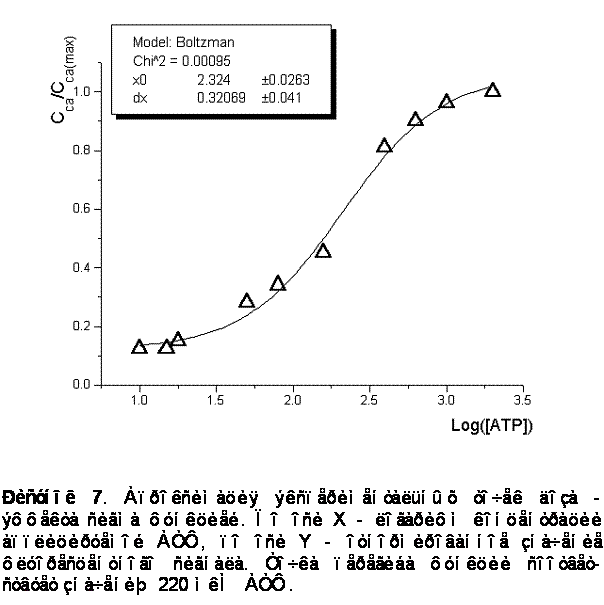
Наблюдалась доза - зависимость амплитуды ответа от концентрации АТФ в пределах от 10 до 2000 мкМ. На рисунке 7 представлена апроксимация экспериментальных точек сигма - функцией. Концентрация АТФ, при которой амплитуда ответа составляет половину максимальной, - 220 ± 20 мкМ.
|
|
Целью наших исследований было определение подтипов пуринорецепторов вовлеченных в генерацию [Ca2+]i транзиента. Известно, что АТФ активирует несколько типов пуринорецепторов, а именно Р2 пуринорецепторы подтипов Р2х и Р2у (Peter Illes et al, 1993, Burnstock and Kennedy 1985), а также Са2+ каналы плазмалеммы (Кришталь, 1983).
С целью обеспечения достоверности полученных данных, принимая во внимание тот факт, что работа проводилась на тонких срезах мозга, мы провели серию экспериментов с подавлением распостранения потенциалов действия с помощью тетродотоксина (TTX). При использовании TTX выяснилось, что амплитуда и форма [Ca2+]i транзиентов не изменяется или изменяется незначительно. Полученные результаты позволяют нам предполагать, что получаемые нами данные отображают процессы происходящие в отдельной наблюдаемой клетке. К сожалению мы не имели возможности проводить все эксперименты c тетродотоксином из-за дороговизны препарата. Для изучения характеристик рассматриваемых пуринорецепторов, принимающих участие в генерации [Ca2+]i транзиента, применялись следующие протоколы экспериментов.
1. Приложение АТФ в растворе не содержащем Ca2+. Удаление кальция из внеклеточного раствора незначительно влияло на кальциевый транзиент, вызванный аппликацией 10 мкМ АТФ, но в тоже время подавляло на 45% ± 7% кальциевый транзиент вызванный 100 мкМ АТФ. (Рис. 8).
Таблица 2. Современная классификация Р2 типа пуринорецепторов.
Исследования проводились на пирамидальных нейронах моторной области новой коры в тонких срезах мозга, выделенного из 14 дневных крыс. После декапитации мозг помещался в холодный солевой раствор (0 - 40С). Процедура от начала декапитации до выделения мозга длилась не более 60-90 секунд. Затем мозг закреплялся полиакриламидным клеем на подложке вибротома (Campden, Campden Instruments LTD, U.K.); камера вибротома заливалась холодным солевым раствором. Срезы нарезались сагитально толщиной 250 - 300 мкм; скорость подачи лезвия - 1 см/с с частотой 10 Гц. После приготовления срезы помещались в раствор постоянно насыщаемый карбогеном (5% СО2 + 95% О2). Перед загрузкой флуоресцентным красителем срезы инкубировались в постоянно оксигенируемом растворе 30 минут при температуре 32 градуса. Окраска среза осуществлялась в течении 30 - 35 минут в СО2 насыщенном термостате при температуре 35 градусов. После окраски срезы отмывались 1 - 1,5 часа в постоянно оксигенируемом растворе при комнатной температуре. Все эксперименты проводились при температуре 32 градуса.
Реферат опубликован: 12/11/2009